
Welcome to the KBLee Group Website!
















Recent News
- Callan won the 2024 Reid Award, Congrats!
- Sarah won the 2024 Van Dyke Award, Congrats!
- Prof. Ki-Bum Lee is accepted into the NJACTS Academy of Mentors, Congrats!
- Li Ling is awarded the NSF Graduate Research Fellowship, Congrats!
- Meizi is awarded the Zhou Fellowship, Congrats!
- The 2023 T32 Immersion Event will be held on January 12th & 13th, 10 am – 5 pm. For more information, including the registration link, see the events tab.
- Skylar won the 2023 Van Dyke Award, Congrats!
- Brandon won the 2023 Reid Award, Congrats!
- Sarah is awarded Biotechnology Training Program Fellowship, Congrats!
- Dr. Jinho Yoon will be joining the Catholic University of Korea (Korea) as an assistant professor this Fall. Congrats!
- Prof. Ki-Bum Lee has been promoted to Distinguished Professor. Congrats!
Prof. Lee would like to acknowledge KBLEE group and collaborators for their support and contributions. - Brian has successfully defended Ph.D. dissertation. Congrats Dr. Conley!
- Brandon is awarded the Training in Translating Neuroscience to Therapies Fellowship, Congrats!
- The 2022 T32 Immersion Event will be held on January 14th-15th, 10 am – 6 pm. For more information, including the registration link, see the events tab.
- Dr. Jin-Ha Choi joined Jeonbuk National University (Korea) as an Assistant Professor. Congrats!
- Prof. Ki-Bum Lee won the 2021 Grossman Innovation Prize, Congrats!
- Brian won the 2020 Reid Award, Congrats!
- Jeff won the 2020 Van Dyke Award, Congrats!
- Hudifah won the 2019 Reid Award, Congrats!
- Chris won the 2018 Reid Award, Congrats!
- Hudifah won the 2018 Van Dyke Award, Congrats!
- Top won the 2018 Van Dyke Award, Congrats!
- Amin won the first annual BME Leadership Award, Congrats!
- Our work on the MnO2 nanoscaffold are now on many news media!
Eurekalert, R&D Magazine, Innovators Magazine, Drug Target Review, The Politico Post, Science Daily, Phys.Org, Innovations Report, Jersey Tribune, News-Medical, Bioengineer, Medical news line, (e) Science News, Front Line Genomics, Science Bulletin, BioSpectrum Asia Edition, W3Live News, BioNews Central, Dove Med, Career Biotech, ChemEurope - Chris won the New Jersey Center for Spinal Cord Research Fellowship, Congrats!
- Skylar is awarded Biotechnology Training Program Fellowship!
- Hudifah, Letao, and Yixiao all won 2018 Professional Development Fund for Teaching and Graduate Assistants!
- Skylar, Joshua, Sarah, Cemile, Hye Kyu, Wankyu, and Hyunjun’s Manuscript published in Advanced Materials: “Enhancing CAR Macrophage Efferocytosis via Surface Engineered Lipid Nanoparticles Targeting LXR Signaling”, Advanced Materials, 2024.
- Euiyeon and Hye Kyu’s Manuscript published in Advanced Functional Materials: “Real-Time, Non-Invasive Monitoring of Neuronal Differentiation Using Intein-Enabled Fluorescence Signal Translocation in Genetically Encoded Stem Cell-Based Biosensors”, Advanced Functional Materials, 2024.
- Letao, Yannan, and Jeff’s Manuscript published in Advanced Materials: “Effective Modulation of Inflammation and Oxidative Stress for Enhanced Regeneration of Intervertebral Discs Using 3D Porous Hybrid Protein Nanoscaffold”, Advanced Materials, 2023.
This is a collaboration work with Prof. Inbo Han at Cha University School of Medicine. - Sarah and Callan’s Manuscript published in Small: “Nanotechnology Approaches to Investigate Chemotherapy-Induced Neurotoxicity, Neuropathy, and Cardiomyopathy in Breast and Ovarian Cancer Survivors”, Small, 2023.
This is a collaboration work with Prof. Mi-Hyeon Jang at Robert Wood Johnson Medical School. - Brian, Letao and Jeff’s Manuscript published in ACS Nano: “Enhanced Intervertebral Disc Repair and Regeneration Using a Nano-Hybrid Peptide Hydrogel”, ACS Nano, 2023.
This is a collaboration work with Prof. Inbo Han at Cha University School of Medicine. - Letao, Chris, Yannan and Brandon’s Manuscript published in Nano Letters: “Nanoparticle-Based Artificial Mitochondrial DNA Transcription Regulator: MitoScript”, Nano Letters, 2023.
- Jeff, Top, Brian, Jinho and Yannan’s Manuscript published in Bioactive Materials: “Injectable BIOorthogonal HydroGEL (BIOGEL) Accelerates Tissue Regeneration in Degenerated Intervertebral Discs”, Bioactive Materials, 2022.
This is a collaboration work with Prof. Inbo Han at Cha University School of Medicine. - Brandon, Brian, Yannan, and Meizi’s Manuscript published in Advanced Drug Delivery Reviews: “Advanced Theragnostics for the Central Nervous System (CNS) and Neurological Disorders Using Functional Inorganic Nanomaterials”, Advanced Drug Delivery Reviews, 2022.
- Letao, Brian, Jinho, Chris, Top, Brandon, and Yannan’s Manuscript published in Research: “High-Content Screening And Analysis of Stem Cell-Derived Neural Interfaces Using Combinatorial Nanotechnology Approach and An Integrated Machine Learning”, Research, 2022.
- Jeff’s Manuscript published in ACS Applied Materials & Interfaces: “Advanced Drug Delivery Via Hybrid Nanofibers Enhance Stem Cell Differentiation”, ACS Applied Materials & Interfaces, 2022.
- Mehrdad and Meizi’s Manuscript published in Journal of Molecular Liquids: “Evaluation of Antibacterial and Mechanical Features of Dental Adhesives Containing Colloidal Gold Nanoparticles”, Journal of Molecular Liquids, 2022.
- Skylar, Brandon, Josh, and George’s Manuscript published in Nano Convergence: “Nanotechnology-enabled immunoengineering approaches to advance therapeutic applications”, Nano Convergence, 2022.
- Jinho and Brian’s Manuscript published in ACS Nano: ” Ultrasensitive Electrochemical Detection of Mutated Viral RNAs with Single Nucleotide Resolution Using a Nanoporous Electrode Array (NPEA)”, ACS Nano, 2022.
This is a collaboration work with Prof. Jeong-Woo Choi at Sogang University. - Letao, Brian, Top, and Brandon’s Manuscript published in ACS Nano: “Predictive Biophysical Cue Mapping for Cellular Differentiation and Reprogramming Using Combinatorial Nanoarrays”, ACS Nano, 2022.
- Letao, Chris, and Yannan’s Manuscript published in Small: “Harnessing the Therapeutic Potential of Extracellular Vesicles for Biomedical Applications using Multifunctional Magnetic Nanomaterials”, Small, 2022.
This is a collaboration work with Prof. Heemin Kang at Korea University. - Jason and Letao’s Manuscript published in National Science Review: “3D Nano-Hybrid Stem Cell Gel for Rapid Cartilage Regeneration”, National Science Review, 2022.
This is a collaboration work with Prof. Qiuyu Zhang at Northwestern Polytechnical University. - Jinho Yoon’s Manuscript published in Small Methods: “Biomolecular Electron Controller Composed of Nanobiohybrid with Electrically Released Complex for Spatiotemporal Control of Neuronal Differentiation”, Small Methods, 2021.
This is a collaboration work with Prof. Jeong-Woo Choi at Sogang University. - Chris and Letao’s Manuscript accepted in Science Advances: “Hybrid SMART Spheroids for Enhanced Stem Cell-based Cell Replacement Therapy”, Science Advances, 2021.
- SangAh, Yixiao, Chris, and Top’s Manuscript published in Advanced Materials: “Bioengineering Approaches for the Advanced Organoid Research”, Advanced Materials, 2021.
- Jin-Ha Choi, Letao, Brian, and Jinho Yoon’s Manuscript published in ACS Nano: “CRISPR-Cas12a-mediated Amplification-free Detection of Multi-Viral DNAs Using SERS-active Graphene Oxide/Triangle Au nanoflower Array””, ACS Nano, 2021.
This is a collaboration work with Prof. Jeong-Woo Choi at Sogang University. - Our Manuscript published in Small: “Magnetic Control of the Vertical Position of Hierarchic Ligand Nanoassembly Regulates the Mechanosensing-Mediated Differentiation of Stem Cells””, Small, 2021.
This is a collaboration work with Prof. Heemin Kang at Korea University. - Collaboration Manuscript with Prof. Heemin Kang published in Advanced Materials: “Remote Control of Time Regulated Stretching of Ligand-Presenting Nanocoils In Situ Regulates Cyclic Adhesion and Differentiation of Stem Cells”, Advanced Materials, 2021.
- Collaboration Manuscript with Prof. Heemin Kang accepted in Advanced Functional Materials: “Remote Switching of Elastic Movement of Decorated Ligand Nanostructures Controls the Adhesion-Regulated Polarization of Host Macrophages”, Advanced Functional Materials, 2021.
- Collaboration Manuscript with Prof. Li Cai published in Experimental Neurology: “Nkx6 Promotes Neurogenesis and Attenuates Glial Scar Formation in the Injured Spinal Cord”, Experimental Neurology, 2021.
- Collaboration Manuscript with Prof. Li Cai published in Molecular Therapy: “Gsx1 Promotes Locomotor Functional Recovery After Spinal Cord Injury”, Molecular Therapy, 2021.
- Collaboration Manuscript with Prof. Morteza Mahmoudi published in Molecular Pharmaceutics: “Restoring endogenous repair mechanisms to heal chronic wounds with a multi-fuctional wound dressing”, Molecular Pharmaceutics, 2021.
- Letao, Brian, and Top’s Manuscript published in Advanced Materials: “Effective Modulation of CNS Inhibitory Microenvironment using Bio-inspired Hybrid Nanoscaffold-based Therapeutic Interventions”, Advanced Materials, 2020.
- Letao, Tae-Hyung, Hyeon-Yeol, Jeff, Jong-Min, Dean, Yannan, and Perry’s Manuscript published in Advanced Functional Materials: “Hybrid Graphene-Gold Nanoparticle-based Nucleic Acid Conjugates for Cancer Specific Multimodal Imaging and Combined Therapeutics“, Advanced Functional Materials, 2020.
- Jeff’s Collaboration Manuscript published in Advanced Materials: “4D Printed Transformable Tube Array for High-throughput 3D Cell Culture and Histology”, Advanced Materials, 2020.
This is a collaboration work with Prof. Howon Lee at Rutgers University - Jin-Ha, Letao, and Brian’s Manuscript published in Nano Letters: “In Situ Detection of Neurotransmitters from Stem Cell-derived Neural Interface at the Single-Cell Level via Graphene-Hybrid SERS Nanobiosensing”, Nano Letters, 2020
- Yixiao and Hudifah’s Manuscript published in ACS Applied Materials & Interfaces: “NIR-mediated Spatiotemporal Control of Stem Cell Fate Using Photo-switching Upconversion Nanoparticle Construct”, ACS Applied Materails & Interfaces, 2020
- Jin-Ho, Jeff, and Dean’s Review Article published in Nanoscale: “Functional nanoarrays for investigating stem cell fate and function”, Nanoscale, 2020
- Letao, Jin-Ho, Chris, and Yannan’s Manuscript published in Nano Letters: “Combinatorial Graphene-Au hybrid SERS nanoarray for Dual-enhanced Neuronal Gene Detection”, Nano Letters, 2019
- Chris’s Review Article accepted in Frontiers in Cellular Neuroscience: “Bio-Inspired Nanomaterials for Delivery of Therapeutics to Treat Hearing Loss”, Frontiers in Cellular Neuroscience, 2019
- Jin-Ho and Jin-Ha’s Manuscript published in ACS Nano: “Nondestructive Characterization of Stem Cell Neurogenesis By a Magneto-Plasmonic Nanomaterial-based Exosomal miRNA Detection”, ACS Nano, 2019
- Jason and Letao’s Manuscript published in Biomaterials: “Programmed Degradation of a Hierarchical Nanoparticle with Redox and Light Responsivity for Self-activated Photo-chemical Enhanced Chemodynamic Therapy”, Biomaterials, 2019
- Hyeon-Yeol’s Collaboration Manuscript is Published in Bioconjugate Chemistry: “Site-specific Incorporation of a Dithiolane Containing Amino Acid in-to Proteins”, Bioconjugate Chemistry, 2019
This is a collaboration work with Prof. Peter Schultz at Scripps - Tae-Hyung, Hyeon-Yeol, and Ahmet‘s Manuscript Published in ACS Applied Materials & Interfaces: “Tumor Homing Reactive Oxygen Species Nanoparticle for Enhanced Cancer Therapy”, ACS Applied Materials & Interfaces, 2019
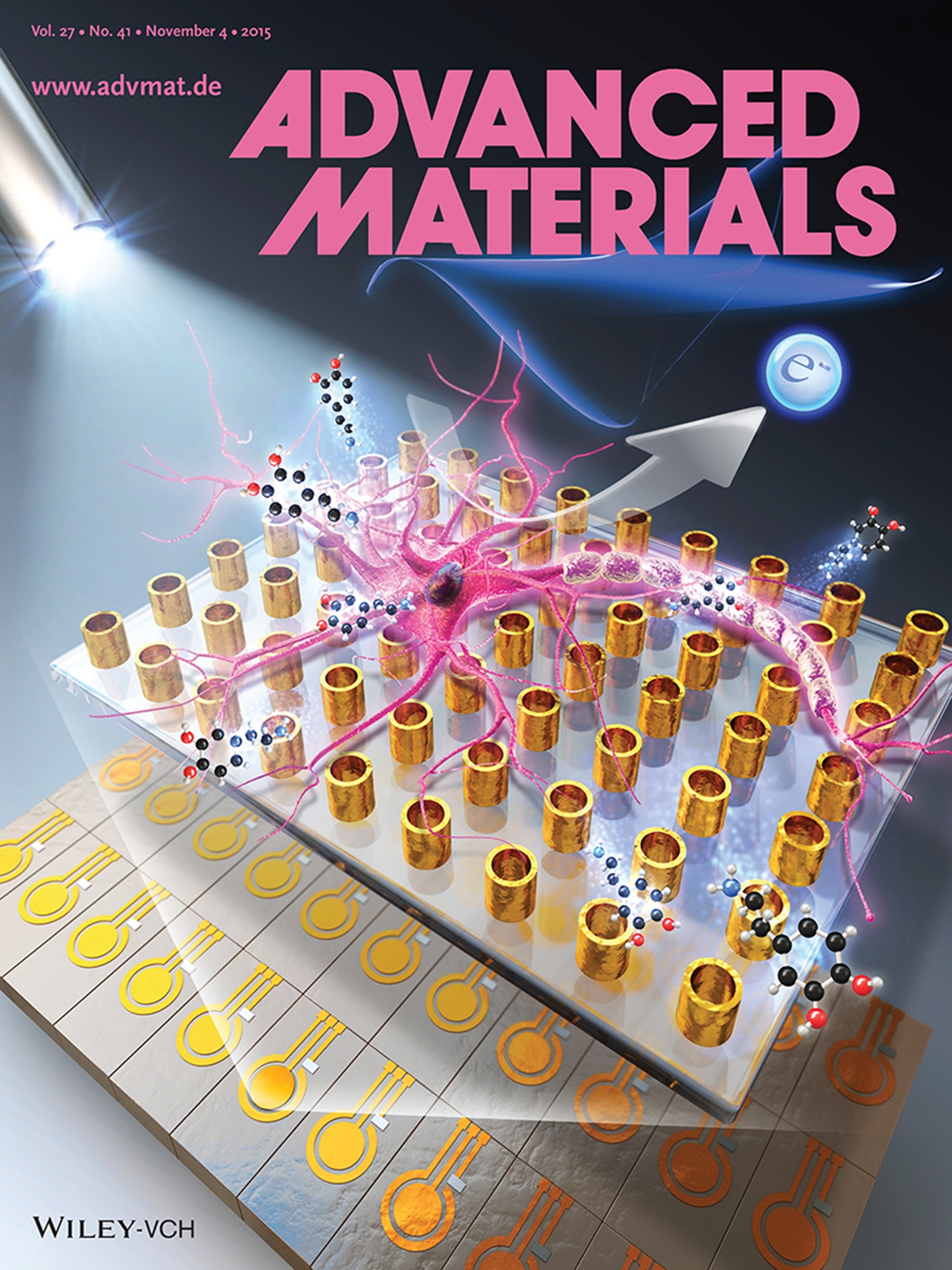
- Hudifah, Yixiao, and Nick’s Manuscript Published in Advanced Materials: “NIR Biosensing of Neurotransmitters in Stem Cell‐Derived Neural Interface Using Advanced Core–Shell Upconversion Nanoparticles”, Advanced Materials, 2019
- Hyeon-Yeol’s Collaboration Manuscript is Published in Nano Letters: “Engineered Mesenchymal Stem Cell/Nanomedicine Spheroid as an Active Drug Delivery Platform for Combinational Glioblastoma Therapy”, Nano Letters, 2019
This is a collaboration work with Prof. Kam Leong at Columbia University - Jong Min and Letao’s Manuscript Published in Scientific Reports: “Generation of uniform-sized multicellular tumor spheroids using hydrogel microwells for advanced drug screening”, Scientific Reports, 2019
This is a collaboration work with Prof. Bong Geun Chung at Sogang University